
 |
| (source) |
It’s only after things got familiarly human in the locomotor anatomy--when we got long legs, non-grasping toes, and reconfigured butts--that brains started increasing beyond ape proportions.
For the first four or five million years of hominin evolution (from 7-2.5 million years ago) the story’s about bipedalism. For the last two and a half, it’s about encephalization. We've known this thanks to fossils for a while and genetic evidence is saying the same thing. It’s natural, then, for such a cerebral organism to wonder whether the two are connected.
As you hypothesize, you could go the technology route. Freed forelimbs, not necessary for locomotion, are free to be handy. O! the possibilities for hurling turds and building worlds! So that's one idea: Selection for a brainier hominin (both physically and cognitively) could occur only after the hands were habitually free to be freaky.
You could go the ecology route. Once our bodies committed to bipedalism our diet changed to include more meat, hominin body size increased, and geographic dispersal did too, no doubt aided by our more efficient bodies built for long distance travel. These characteristics, together, have been compared to those of scavenging and predatory carnivores. Regardless of how small or large a part meat played in our ancestors’ diets, there’s no denying that an ecological shift occurred in the early Pleistocene, with an increase in diet and habitat diversity, and that shift must have included new requirements of the brain.
Or, you could go the sociality route. As hominins relied more and more on cooperative foraging and parenting behaviors, etc, navigating social networks became key. Once complex speech and language arrived, then there would be new demands on the brain as well.
These pressures, requirements, demands, however you want to think of them, could be working in concert and at different times (e.g. technology plus socializing) over deep, geologic time and many many hominin generations. By "working," I mean contributing to the more-or-less sustained differential reproductive success of hominins with slightly larger brains. And because it’s the way that the fossil and archaeological records reveal behaviors over time, I tend to think of these three categories (technology, ecology, sociality) as describing the last 2.5 million years in the order I listed them. Technology was strongest earliest (starting with the Oldowan stone tools by 2.6 mya), and persisted. An ecological shift came along with that technological shift and then persisted. And of course social complexity came along with the technological and ecological shifts and then persisted.
These are some of the most mainstream hypotheses for encephalization (1) and they're implicitly or explicitly predicated on the prior evolution of bipedalism.
But now there's a new tie between big brains and walking upright--offered up in a new paper just out in PNAS--and it's based in the human-, not ape-, like tendency to fuse the metopic suture later in life, to delay the close of what starts as a hole in the top of a baby's head known as its anterior fontanelle. The authors suggest that we need this hole in our head to exit our mother's bipedally-adapted birth canal safely and we also, as they suggest, need it to grow up to be an encephalized creature.
According to the authors, the "Taung child", an Australopithecus africanus kid (a member of a well-known bipedal hominin lineage) had an unfused metopic suture, left as an imprint on the fossil brain endocast.
 |
| Raymond Dart with the Taung child fossil. http://en.wikipedia.org/wiki/File:Raymond_Dart_with_Taung.jpg |
{kind=link}
However, to interpret the Taung child’s anatomy, things get a bit dicey, like things just love to get with hominin fossils. So often they can go either way: chimpy or humany.
Check out the figures below. A is for Pan troglodytes (common chimps) and B is for Homo sapiens. Those are frequencies of metopic suture fusion per dental age group. By listing them this way, instead of by chronological age, we're able to compare between two species that grow at different rates but share the same pattern of dental eruption. Chimpanzees grow up faster than us and experience earlier metopic suture fusion than us. Flipped around, humans grow up slower than chimps and experience delayed fusion of this suture compared to them. The Taung baby's dental age category is starred (*) at the "M1" stage in A and B. (The Taung child died at around 3.8 years of age, when its first permanent molar, M1, was erupted.)

"The presence of a still patent fontanelle and of a partially fused [metopic suture] in the Taung child, and the incidence of unfused [metopic sutures] in five adult and two other younger Australopithecus/ early Homo specimens is thus taken as evidence that a human-like pattern of late [metopic suture] fusion was already present in mid-to-late Pliocene gracile hominins."
Okay. Intriguing! But now we must explain!
[This is the part where, if you listen very carefully, you can hear the collective curmudgeonly groans from within and beyond the walls of paleoanthropology.]
Enter the new hypothesis for encephalization based on the late fusion of metopic sutures. The authors nod to two papers that offer "adaptively neutral" explanations for late metopic suture fusion but argue that the fossil evidence combined with the differences observed in chimps and humans beg for an adaptive explanation. (This tack is unsurprising given how paleoanthropology generally operates.)
The authors offer us three adaptive hypotheses to explain late metopic suture fusion:
Something about the changing and enlarging frontal cortex required changes to the cranial bones, how they form, grow, and fuse.
2. The difficulty of giving birth to large-headed neonates through birth canals that were reconfigured for bipedalism, the “obstetrical dilemma” (explains late metopic suture fusion)
The squishy neonatal head, thanks to the fontanelle, "probably occurred in conjunction with refining the ability to walk on two legs," Falk (the lead author) said to the media. "The ability to walk upright caused an obstetric dilemma. Childbirth became more difficult because the shape of the birth canal became constricted while the size of the brain increased. The persistent metopic suture contributes to an evolutionary solution to this dilemma."
The trouble with this hypothesis as applied here is, although we know modern humans have a tight fit at birth now, there's little evidence for a tight fit between neonate and birth canal during australopith times.
And you can't help but wonder whether a squishy head was, or still is, required for successful birth. Do children suffering from craniosynostosis require a c-section to be born? Also, since the metopic suture fuses after chimpanzee birth, are we certain they aren't squishing their brains as they exit their relatively roomy birth canals? These questions may sound silly, but they're illustrating the built-in assumptions of the paper (or my ignorance about squishiness of baby heads).
The squishy head may be helpful during childbirth, but if it's occuring as early as australopith times, an adaptive explanation as a "solution" to an obstetrical dilemma is hard to swallow. That is unless DeSilva's estimates cited by the authors-- that australopiths had large neonates and tight fits at birth--are correct.
3. High early postnatal brain growth rates (explains late metopic suture fusion)
We know that humans have high rates of postnatal brain growth and this is what a lot of the news media picked up on: Your baby's head has gaps between the bones so the brain can grow like crazy after it's born to the gargantuan size of an adult human brain. As established in hypothesis #2, the need for the fontanelles in the first place is the crunch at birth thanks to the obstetrical dilemma, implying that without the pressure to be born small enough to escape the bipedal birth canal, we'd grow larger fetal brains in the womb.
So with this new paper we're presented with something even more fundamental than the notion that bipedalism as a necessary precursor for technological, ecological and social selection pressures for encephalization (as covered above): The tight fit at birth, caused by antagonistic selection for bipedal pelvic anatomy and large neonatal brains, created the selection pressure for a squishy neonatal head (which is facilitated by the fontanelles) and because of that roomy cranium, postnatal growth rates were able to ramp up in selective environments that favored encephalization.
So I'm left wondering, Do we need a hole in our head to be born successfully? Do we need a hole in our head to be encephalized? If the answer to both of those is yes, then what is a hole in the head doing in a hominin genus that may not have had much difficulty with childbirth and was hardly (if at all) encephalized? And, given the overlapping chimp and human fusion patterns, how can we be sure this feature on Taung is humany and not chimpy?
And, further, you can't (or at least I can't) help but wonder if there's a biomechanical/functional explanation for late metopic suture fusion, given how feeding behaviors and masticatory muscles put stress on the cranium. The skulls of australopiths and other hominins experienced stresses differently than chimpanzees. These differences may have begun as early as the nursing stage. Could this have anything to do with delayed fusion of the sutures? (here's just one study I found that addresses these kinds of questions)
And finally, it's hard not to link Falk (the lead author) to her research on Homo floresiensis. The hobbit (LB 1) looks like it has a fontanelle, something the disease-hypothesis folks point out is consistent with their perspective, and that’s one reason why I assumed these authors are onto this topic.
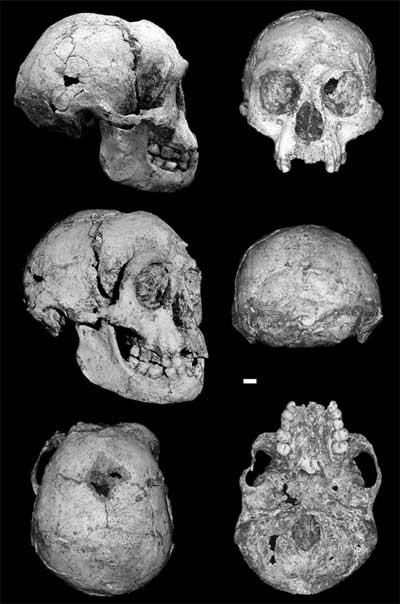
Good thing, because if the hole in LB 1's cranium is of biological and not of taphonomic origin, then who knows how anybody'd explain its adaptive significance in such a tiny-brained hominin.
But, going way back to Taung and the australopiths: They were, after all, bipedal and the big brain train had to pull out at some point!
And, stay tuned. I got a tip from one of the authors about a paper coming out soon that demonstrates how weak the obstetrical dilemma hypothesis is, for explaining fetal size and growth, given the current evidence and given what we know about maternal metabolism.
Notes
(1) Of course, these hypotheses don’t represent all of paleoanthropology. I just intended to cover the major bases. And you need to consider what many paleoanthropologists assume which is that brain tissue is expensive so something extraordinary must have kept up selection on its increasing size for the last 2.5 million years. The assumption is that if brains were cheap, everyone would have big ones, but I don’t buy that. I think it's clear that other species aren’t encephalized because they don’t have to be. They do just fine without big brains. We have a rather warped perspective on selection for encephalization, thanks to our presentism and our big brains.
(2) Thanks to K. Baab for the tip.
8 comments:
As usual, a very fine consideration of this latest adaptive interpretation of human fossil remains. The problem, goes beyond the juicy temptation to assert some specific adaptive mechanism--an open field for speculation as the record of that clearly shows.
The problem is that if you assume adaptation (that is, that the trait is really a trait and it really was affected by natural selection), then you are clearly not constrained by the facts to a particular interpretation.
And if you are more circumspect about selective determinism, then there is no good way to prove neutrality--that, in some way, the trait existed not because of an adaptive force at all, or that it reflects correlated evolution related to some other trait affected by development and/or the same set of gene(s).
And finally, the reward system favors not being very circumspect. This is why many people, largely correctly I'm sad to say, feel that 'anthropology is just story-telling.'
My point can perhaps be made another way. What about the 'aquatic ape' hypothesis? If we evolved 'to' swim in Lake Victoria, does having a big head make swimming easier, and hence could that have demanded the metopic suture in some way?
Or, to invoke Dean Falk's own earlier hypothesis,the 'radiator hypothesis', does a big head owe its evolution to head dissipation, rather than any of your Big 3 traditional reasons (I haven't read the PNAS paper yet, so don't know if Dean addressed this).
I'm hoping that people can read both meanings of my title. I worked so hard on it! Dedicated to all the grammar lovers out there.
Anyone who can't see that clever pun has...a hole in their head!
RE: the hole in the head of LB1, check out Kaifu et al.'s recent paper in the J. Human Evol. It's a detailed analysis of the cranium, and it has micro-CTs of the bregmatic region in question (which Brown et al. [2004 Nature] noted was damaged during the excavation).
BTW, do you buy DeSilva's PNAS argument?
Thanks, I will when I'm back on campus with access. Is it contra Brown? Is that why you mention it?
About DeSilva's PNAS paper on large australopith neonates... I'm having trouble understanding the methods and their implications well enough to have an opinion other than my high opinion of Jerry as a conscientious scientist. I tend, as we all should, to be skeptical about models that are based on so many assumptions and unknowns.
I have found this blog post really helpful thank you. I have been researching this topic for my work, for which I am trying to write a non-technical review of how bipedalism and encephalisation are linked. (I am not an academic nor an expert in this field: a few courses in vertebrate and mammalian evolution at university 11 years ago is all I have to base my understandings on.) I have been coming up with exactly the sorts of questions you have so eloquently described here, finding current explanations incomplete. In particular I am wondering about the supposed advantages of fontanelles during birth. It is often implied that fontanelles evolved to ease the difficulty of birth through the bipedal pelvis (I am aware of the recent paper about energetics/metabolism driving timing of birth but that paper still asserts that human birth is difficult relative to primate relatives because of the narrow hip/large head issue). If that were the case though, why do chimps have fontanelles? I have read that these fuse soon after birth but I haven't been able to find out whether, at birth (before MS fusion occurs) the fontanelles allow the chimp baby's head to 'squish' to ease birth. If not, what is it about human baby fontanelles/skulls that is 'special' that it allows them to 'squish' - and when did that evolve?
I’d be really interested to know if you have found out anything more about this since your blog post. Thanks!
Long story short: perhaps we are too aware of how degreed, rather than unique, too many of our human traits are and this is making it impossible to tell a step by step story of hominin evolutionary history, as if each trait arose in its own time for its special reason, as was (and still can be) the habit of paleoanthropology-thinkers to do.
If you email me, holly_dunsworth@uri.edu, I'll send you a pdf of a recent paper in PNAS that contests the one I blogged about in this post. It's by Ralph Holloway, Doug Broadfield and Kris Carlson. That is, if you don't have access: http://www.pnas.org/content/111/36/13022.full.pdf
Post a Comment